
“Pronađen je lijek koji pobjeđuje virus.” Današnje izdanje “Il Fatto Quotidiano” započinje citatom. “Pronađeno antititijelo koje spriječava zarazu korona virusa!”
La Gismondo: “To je prva dobra vijest, mogli bimo ga iskoristiti za koji mjesec!” Iz Nizozemske dolazi lijek: Antitijelo pobjeđuje Covid.” Zove se “47D11”: Identificirali su ga znanstvenici iz Utrechta i Rotterdama.
Istraživanje je na web stranici BioRxiv objavila skupina sa nizozemskog sveučilišta u Utrechtu koju je vodio Chunyan Wang.
Lijek je monoklonsko antitijelo, specijalizirano za prepoznavanje proteina koji virus koristi za napad ljudskih respiratornih stanica. No, proći će nekoliko mjeseci prije nego što lijek bude dostupan jer nije prošao testiranje.
Znanstvenici su već radili na antitijelu protiv SARS-a kada je eksplodirala epidemija Covid-19. Brzo su shvatili da stvorena antitijela mogu biti djelotvorna protiv Korona virusa.
Vezujući se za protein spike, koji se nalazi na površini koronavirusa, monoklonsko antitijelo “47D11” sprečava vezanje na stanice i na taj način onemogućuje virusu prodiranje unutar njih te tako zaustavi razmnožavanje. Iz tog razloga, istraživači su uvjereni da antitijelo ima važan potencijal “za liječenje i prevenciju Covida 19”.
Linkovi:
Znanstveni tekst pročitajte u nastavku.
Ovaj članak nije ovjeren od strane stručnih recenzenata.

Ljudsko monoklonsko antitijelo koje blokira SARS-CoV-2 infekciju
Sažetak
Pojava novog humanog koronavirusa SARS-CoV-2 u Wuhanu u Kini uzrokovala je svjetsku epidemiju respiratornih bolesti (COVID-19). Trenutno nema cjepiva i ciljanih terapija za liječenje ove bolesti. Ovdje izvješćujemo o ljudskom monoklonalnom antitijelu koje neutralizira SARS-CoV-2 (i SARS-CoV). Ovo unakrsno neutraliziranje antitijela cilja zajednički epitop na ove viruse i nudi potencijal za prevenciju i liječenje COVID-19.
Glavni tekst
Teški akutni respiratorni sindrom koronavirus 2 (SARS-CoV-2) etiološki je uzročnik bolesti izazvane koronavirusom (COVID-19) koja se pojavila u Kini krajem 2019. i uzrokovala svjetsku epidemiju 1 . Od 10. ožujka 2020. u 109 zemalja prijavljeno je preko 115.113 slučajeva, od kojih je 4.063 (3,5%) podleglo infekciji 2 . SARS-CoV-2 pripada podgrupi Sarbecovirus (rod Betacoronavirus , obitelj Coronaviridae ) 3zajedno s SARS-CoV-om koji se pojavio 2002. godine uzrokujući oko 8000 infekcija sa smrtnošću od 10%. Oba virusa prelaze barijere vrsta iz životinjskog rezervoara i mogu uzrokovati po život opasne respiratorne bolesti kod ljudi. Trenutno nisu dostupni odobreni ciljani terapeutici za COVID-19. Monoklonska antitijela koja ciljaju ranjiva mjesta na virusnim površinskim proteinima sve su više prepoznata kao obećavajuća klasa lijekova protiv zaraznih bolesti i pokazala su terapijsku učinkovitost za brojne viruse 4 , 5 .
Antitijela koja neutraliziraju koronavirus primarno ciljaju glikoproteine trimernog spike (S) na površini virusa koji posreduju ulasku u stanice domaćina. S protein ima dvije funkcionalne podjedinice koje posreduju vezivanje stanica (S1 podjedinica, postojeće od četiri jezgrene domene S1 A do S1 D ) i fuziju virusne i stanične membrane (S2 podjedinica). Moćna protutijela za neutralizaciju često ciljaju mjesto interakcije receptora u S1, onemogućujući interakcije receptora 6 – 11 . Proteini šiljaka SARS-CoV-2 (SARS2-S; 1.273 ostatka, soj Wuhan-Hu-1) i SARS-CoV (SARS-S, 1.255 ostataka, soj Urbani) sa 77.5% su identični po sekvenci primarne aminokiseline, strukturno vrlo slično 12 , 13i obično se vežu na protein enzima 2 (ACE2) koji pokriva humani angiotenzin kao domaćin receptora 1 , 14 kroz S1 B domenu. Poznato je da interakcija receptora pokreće nepovratne konformacijske promjene u proteinima spike koronavirusa što omogućava fuziju membrane 15 .
Kako bi se identificirala protutijela koja neutraliziraju SARS-CoV-2, procijenjena je ELISA- (unakrsna) reaktivnost supernanata koji sadrže antitijela iz kolekcije 51 SARS-S hibridoma dobivenih od imuniziranih transgenih miševa H2L2 koji kodiraju himerne imunoglobuline s promjenjivim ljudskim teškim i lakim lanci i konstantna područja podrijetla štakora ( Suppl.Fig.1 ). Četiri od 51 supernatanta SARS-S hibridoma pokazali su ELISA-cross-reaktivnost sa SARS2-S1 podjedinicom (S ostaci 1-681; Suppl.Fig.1 ), od kojih je jedan (47D11) pokazao aktivnost ne-neutraliziranja SARS-S i SARS2-S pseudotipiziranom VSV infekcijom. Himerno 47D11 H2L2 antitijelo je preoblikovano i rekombinantno eksprimirano kao potpuno ljudsko IgG1 izotipilo za daljnje karakteriziranje.

Suppl. Sl. 1.ELISA unakrsna reaktivnost supernanata koji sadrže antitijela SARS-S H2L2 hibridoma prema SARS2-S1.
SARS-S ciljani hibridomi razvijeni konvencionalnom tehnologijom hibridoma iz imuniziranih H2L2 transgeničnih miševa (Harbor Biomed), kako je opisano prije 1 . Ti su miševi – koji nose kodiranje repertoara imunoglobulina teškog i lakog lanca – bili sekvencijalno imunizirani s dvotjednim intervalima trimetarskim šiljastim ektodomainima (S ecto ) tri ljudska koronavirusa iz roda betakoronavirusa sljedećim redoslijedom: 1. HCoV-OC43- S ecto , 2. SARS-CoV-S ecto , 3. MERS-CoV-S ecto , 4. HCoV-OC43-S ecto , 5. SARS-CoV-S ecto , 6. MERS-CoV-S ecto, Četiri dana nakon zadnje imunizacije sakupljeni su splenociti i limfni limfni čvorovi i stvoreni su hibridomi. Antitijela u staničnim supernatantima testirana su na ELISA-reaktivnost protiv SARS-S ecto , SARS-S1, SARS-S1 A i SARS2-S1. Od 51 hibridoma supernatanta koji su reagirali samo sa SARS-S ektorom , 23 je reagirao sa SARS-S1 A , 22 sa SARS-S1, ali ne SARS-S1 A , 6 sa SARS-S ekto, ali ne i SARS-Sl. Četiri od 51 supernatanta ekto hibridoma SARS-S reagirali su sa SARS2-S1 (vidi stupac s desne strane). Tablica prikazuje intenzitete ELISA signala ( vrijednosti OD 450nm ) hibridomskih supernanata za različite antigene.
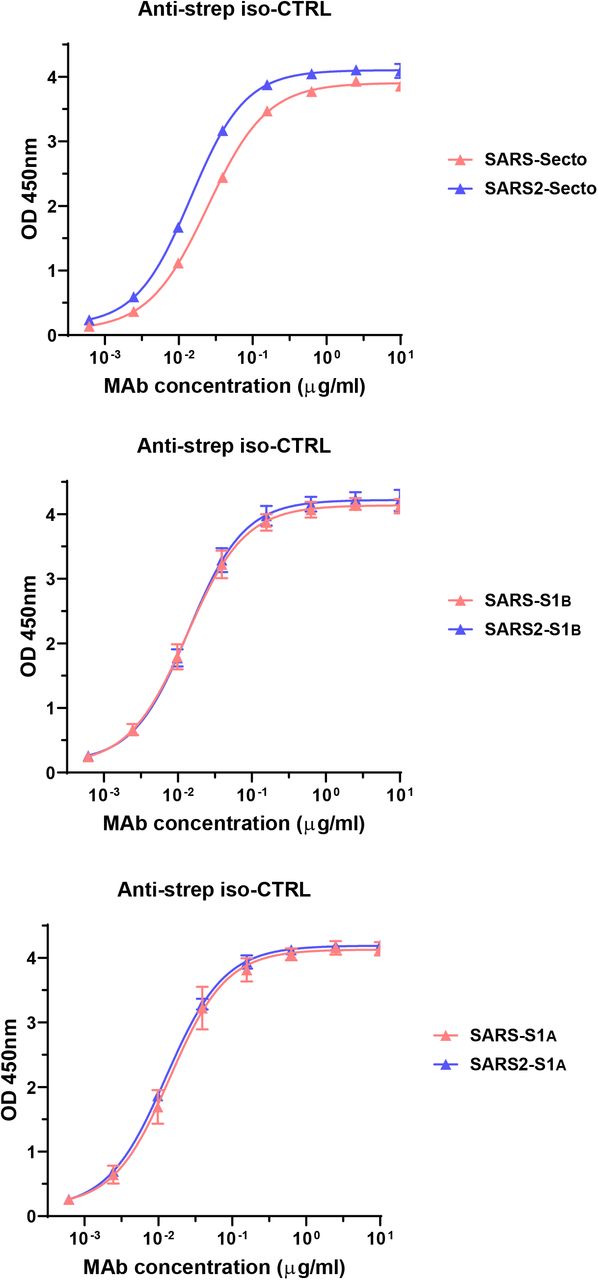
Humana 47D11 antitijela se vežu na stanice koje eksprimiraju proteine spike pune duljine SARS-CoV i SARS-CoV-2 ( Sl.1a ). 47D11 protutijelo pokazalo je potencijalno inhibiraju infekcije stanica sa VeroE6 SARS-S i S-SARS2 pseudotyped VSV s IC 50 vrijednost 0,06 i 0,08 | ig / ml ( Fig.1b ), respektivno. Autentični infekcije stanica sa VeroE6 SARS-COV i SARS-COV-2 je neutralizirana IC 50 vrijednostima 0.19 i 0.57 ug / ml ( Slika 1C ). Korištenje ELISA 47D11 pokazalo je ciljanje domene vezivanja S1 B receptora (RBD) SARS-S i SARS2-S. 47D11 vezao je S1 B oba virusa sa sličnim afinitetima, kao što pokazuje polovica maksimalne učinkovite koncentracije temeljene na ELISA (EC50 ) vrijednosti (0,02 i 0,03 µg / ml, respektivno; Sl.2a ). ELISA afinitet vezanja 47D11 za šiljak ektodomensko (S ecto ) SARS-COV bila veća u odnosu na onaj SARS-COV-2 (EC 50 vrijednost: 0.018 i 0.15 ug / ml, respektivno), bez obzira na premaz ekvimolarnom antigena ( Suppl.Fig.2 ). Istodobno s ELISA reaktivnošću, mjerenje kinetike vezivanja 47D11 biološkom interferometrijom pokazalo je da 47D11 veže SARS-S ekto s većim afinitetom (konstanta ravnotežne disocijacije [ K D ]: 0.745 nM) u odnosu na SARS2-S ekto ( K D 10.8 nM ) budući da je afinitet za SARS-S1 B i SARS2-S1B je bio u sličnom rasponu (16,1, odnosno 9,6 nM, Suppl.Fig.3 ). Ova razlika može potjecati od razlike epitop dostupnosti u SARS-S odnosu SARS2-S, kao i domene B može usvojiti zatvorenu i otvorenu strukturu u prefusion šiljka homotrimer 12 , 13 . Izvanredno, vezanje za 47D11 SARS-S1 B i SARS2-S1 B ne natječu s S1 B vezanja na receptor izražen ACE2 na površini stanice, kao što je prikazano pomoću protočne citometrije ( Fig.2b ; Suppl.Fig.4 ) niti u S ekto i S1 B vezanje za topljivi ACE2 u testu zasnovanom na krutoj fazi ( Suppl.Fig.5), dok dva SARS-S1 specifična antitijela 35F4 i 43C6 koja neutraliziraju SARS-S (ali ne i SARS2-S) pseudotipiziranu VSV infekciju ( Suppl.Fig.6 ) čine blokadno vezivanje SARS-S ecto i SARS-S1 B na ACE2. Korištenjem testa staničnih stanica aktiviranog tripsinom, pokazalo se da 47D11 smanjuje stvaranje sincicija posredovanih SARS-S i SARS2-S ( Suppl.Fig.7 ). Naši podaci pokazuju da 47D11 neutralizira SARS-CoV i SARS-CoV-2 putem još nepoznatog mehanizma koji se razlikuje od smetnji vezivanja receptora. Zabilježeni su alternativni mehanizmi neutralizacije koronavirusa pomoću antitijela usmjerenih na RBD, uključujući inaktivaciju šiljaka kroz destabilizaciju njegove predfuzione strukture 15 izazvane protutijelom , koja se također može primijeniti na 47D11.

Suppl. Sl.2.
ELISA krivulja vezanja anti-StrepMAb (IBA), antitijela na Strep-markirane klasa antigena da potvrdi ekvimolarnom ELISA ploče prevlačenje SARS-S ecto / SARS2-S ecto (gornji panel), SARS-S1 B / SARS2-S1 B (srednja ploča) i SARS-S1 A / SARS2-S1 A (donji panel) antigeni korišteni na Sl.2a .
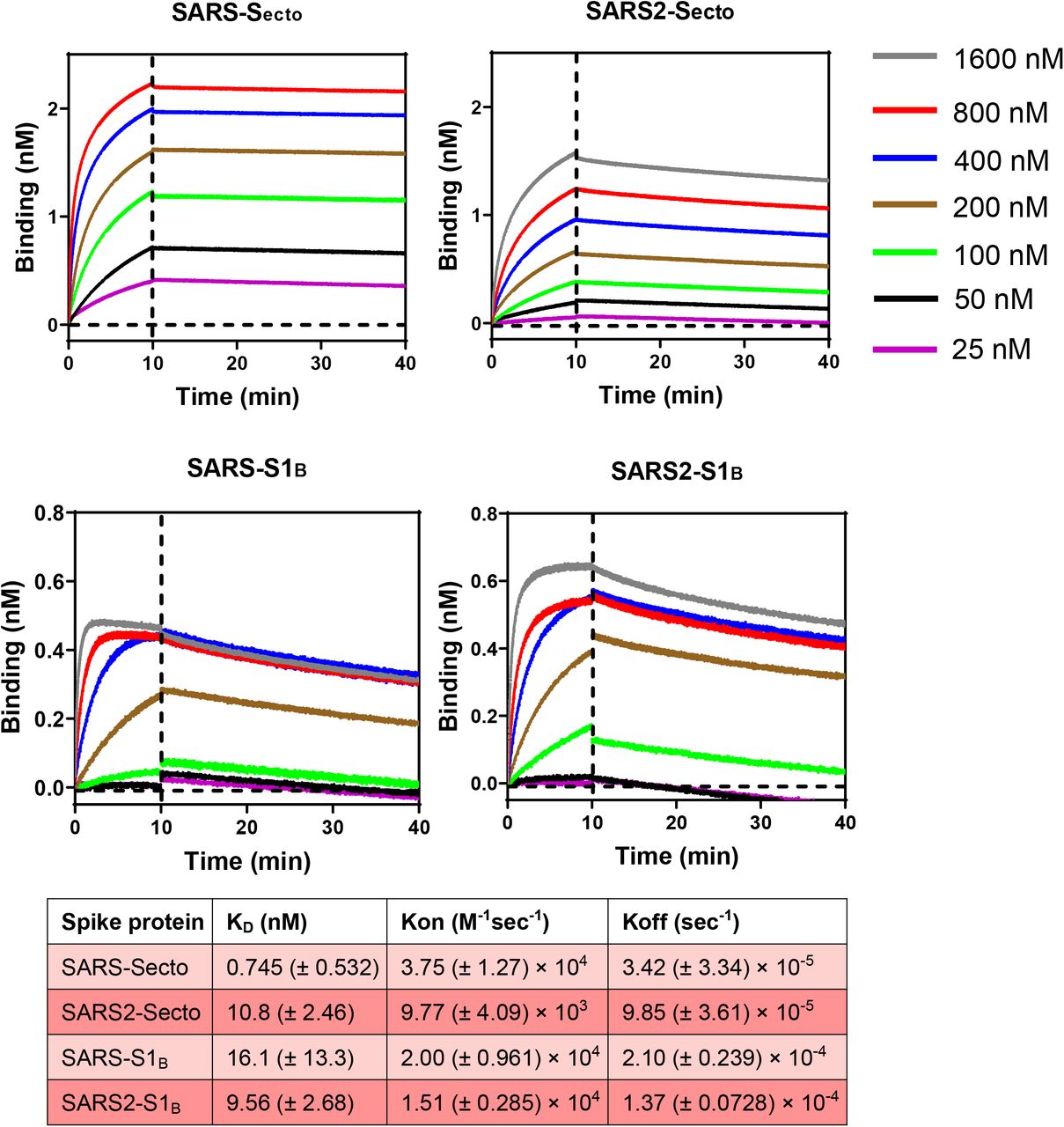
Suppl. Slika 3.Kinetika vezanja 47D11 na S ektodomenu i S1 B SARS-CoV i SARS-CoV-2.
Kinetika vezanja na imobilizirani rekombinantni 47D11 SARS-S ecto , SARS2-S ecto , SARS-S1 B i SARS2-S1 B mjerena je pomoću biolayer interferometara na 25 ° C, kao što je prethodno opisano 21 . Ispitivanje kinetičkog vezanja izvedeno je punjenjem 47D11 mAb u optimalnoj koncentraciji (42 nM) na biosenzor protiv humanog Fc u trajanju od 10 minuta. Korak povezivanja antigena izveden je inkubacijom senzora s rasponom koncentracija rekombinantnog ektodomaina šiljka (1600-800-400-200-100-50-25 nM) tijekom 10 minuta, nakon čega slijedi korak disocijacije u PBS-u 60 minuta. Konstante kinetike izračunane su korištenjem 1: 1 Langmuirovog modela vezivanja na Fortebio Data Analysis 7.0 softveru.

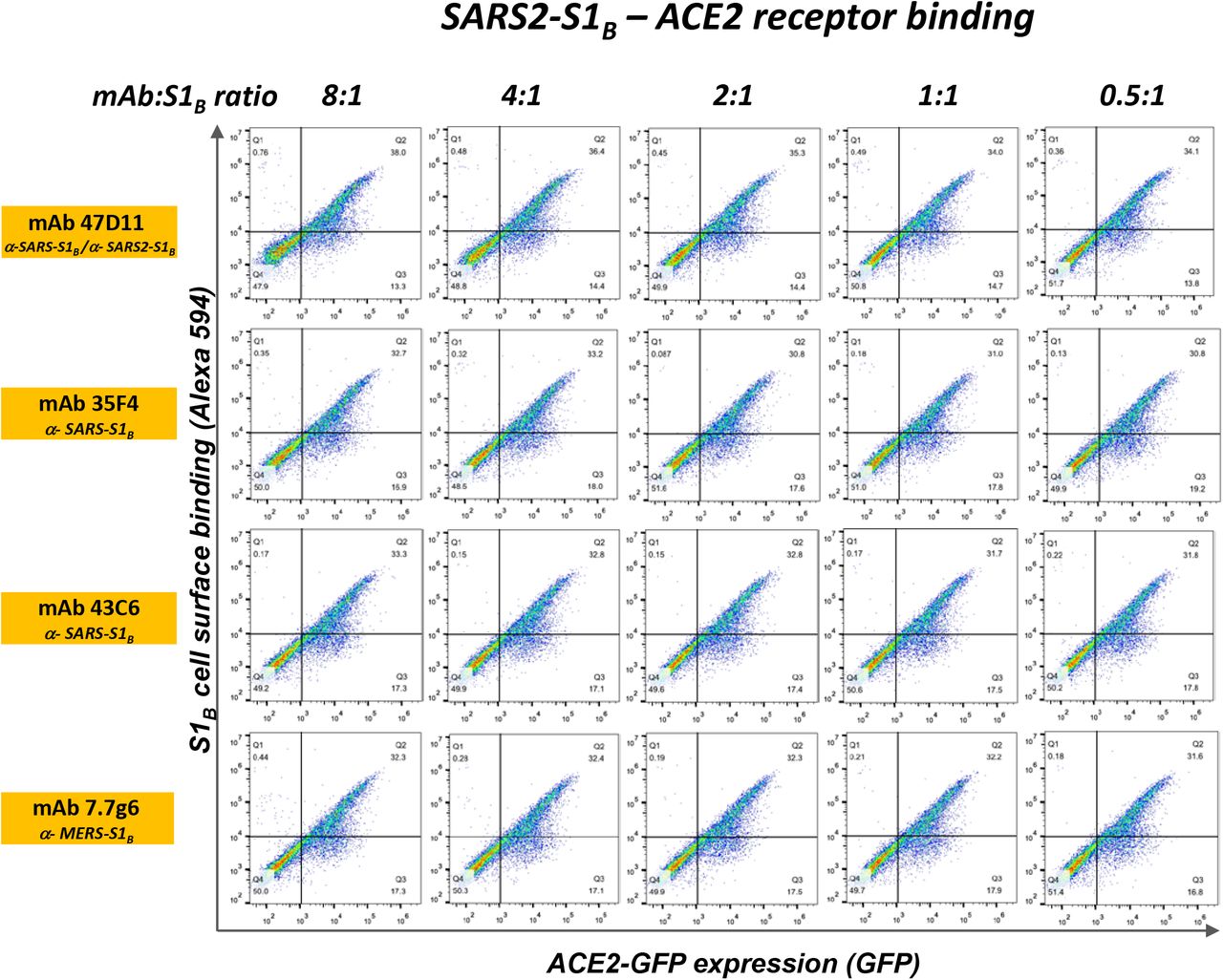
Suppl. Slika 4.47D11 ne sprječava vezanje SARS-S1 B i SARS2-S1 B u ACE2-ekspresijskih stanica.
Ljudske HEK-293T stanice koje eksprimiraju humane ACE2-GFP proteine (vidjeti metode) odvojile su se i fiksirale s 2% PFA, inkubirale s fiksnom količinom humane Fc označene S1 B domene SARS-S ili SARS2-S koja je prethodno inkubirana 1 sat s mAb (mAbs 47D11, 35F4, 43C6, 7.7G6, u H2L2 formatu) u naznačenim molarnim omjerima mAb: S1 B , i analizirano protočnom citometrijom pomoću sekundarnog antitijela povezanog s Alexa Flour 594 usmjerenim na ljudsku Fc oznaku. Stanice se analiziraju na GFP ekspresiju (x-os, GFP signal) i vezivanje antitijela (y-osi, signal Alexa 594). Postoci ćelija s negativnim, pojedinačnim ili dvostrukim pozitivnim vrijednostima prikazani su u svakom kvadrantu. Kontrole vezivanja uključuju stanice tretirane PBS-om (izravnavanje), liječenje stanica s SARS-S1 B i SARS2-S1 Bu nedostatku antitijela i stanica liječenih samo antitijelima. Eksperiment je izveden dva puta, prikazani su podaci reprezentativnog eksperimenta.

Suppl. Slika 5.Test inhibicije vezivanja receptora na bazi ELISA.
Test inhibicije vezivanja receptora na bazi ELISA izveden je kao što je prethodno opisano s nekim prilagodbama 1 . Rekombinantni topljivi ljudski ACE2 preko noći je obložen na NUNC Maxisorp pločama (Thermo Scientific) na 4 ° C. Ploče su isprane tri puta s PBS-om koji je sadržavao 0,05% Tween-20 i blokiran s 3% BSA u PBS-u koji je sadržavao 0,1% Tween-20 na sobnoj temperaturi tijekom 2 sata. Rekombinantni S ecto i S1 B SARS-S ili SARS2-S (300 ng) i serijski razrijeđeni mAbs (mAbs 47D11, 35F4, 43C6, 7.7G6, u H2L2 formatu) miješani su 1 sat na RT, dodani u ploču za Sobna temperatura 1 t, nakon čega su ploče oprane tri puta. Vezivanje na ACE2 detektirano je korištenjem StrepMAb-a konjugiranog HRP-om (IBA) koji prepoznaje C-terminalni streptag na S ecto-ui S1 B proteina.

Suppl. Sl.6.
H2L2 monoklonska antitijela 35F4 i 43C6 neutraliziraju SARS-CoV, ali ne i SARS-CoV-2. Antitijela posredovana neutralizacijom infekcije VSV česticama pseudotipiziranih sa šiljastim proteinima SARS-CoV (gornja ploča) i SARS-CoV-2 (donja ploča) pomoću antitijela 35F4 i 43C6 H2L2 koja ciljaju SARS-S1, ali ne i SARS2-S1 (vidi Suppl .Fig.1 ). Irelevantno antitijelo uzeto je kao humana IgG1 izotipa kontrola. Prikazana su sredstva ± SD trostrukih primjeraka.

Suppl. Slika 7.Test inhibicije stanične stanice fuzije.
Ispitivanje inhibicije stanične stanice-fuzije provedeno je kao što je prethodno opisano s nekim adaptacijama 1 . Stavljene su VeroE6 stanice s gustoćom od 10 5 stanica po ml. Nakon postizanja konfluencije od 70 do 80%, stanice su transfektirane plazmidima koji kodiraju SARS-S, SARS2-S i MERS-S-C koji su krajnje spojeni s GFP-om koristeći Lipofectamine 2000 (Invitrogen). Mjesto prepoznavanja furina u SARS2-S bilo je mutirano (R 682 RAR do A 682AAR) inhibirati cijepanje proteina endogenim furinom i omogućiti stvaranje sincicije uzrokovano tripsinom. Dva dana nakon transfekcije, stanice su prethodno obrađene samo DMEM ili DMEM s 20 µg / ml mAbs tijekom 1 sata, a potom su tretirane DMEM s 15 µg / ml tripsina (za aktiviranje fuzijske funkcije fuzije) u odsutnosti ili prisutnosti 20 µg / ml mAbs (47D11 crossreactive SARS-S i SARS2-S, 35F4 reaktivan na SARS-S, 7.7G6 reaktivan na MERS-S). Nakon inkubacije na 37 ° C tijekom 2 sata, stanice su fiksirane s 2% PFA u PBS-u 20 minuta na sobnoj temperaturi i obojene za jezgre sa 4,6-diamidino-2-fenilindol (DAPI). Stanice koje eksprimiraju S-GFP proteine detektirane su fluorescentnom mikroskopijom i opažena je S-posredovana fuzija staničnih stanica formiranjem fluorescentne) multi-nukleirane sincitije. Fluorescentne slike zabilježene su upotrebom Leica SpeII konfokalnog mikroskopa. Eksperiment je izveden dva puta, prikazani su podaci reprezentativnog eksperimenta.
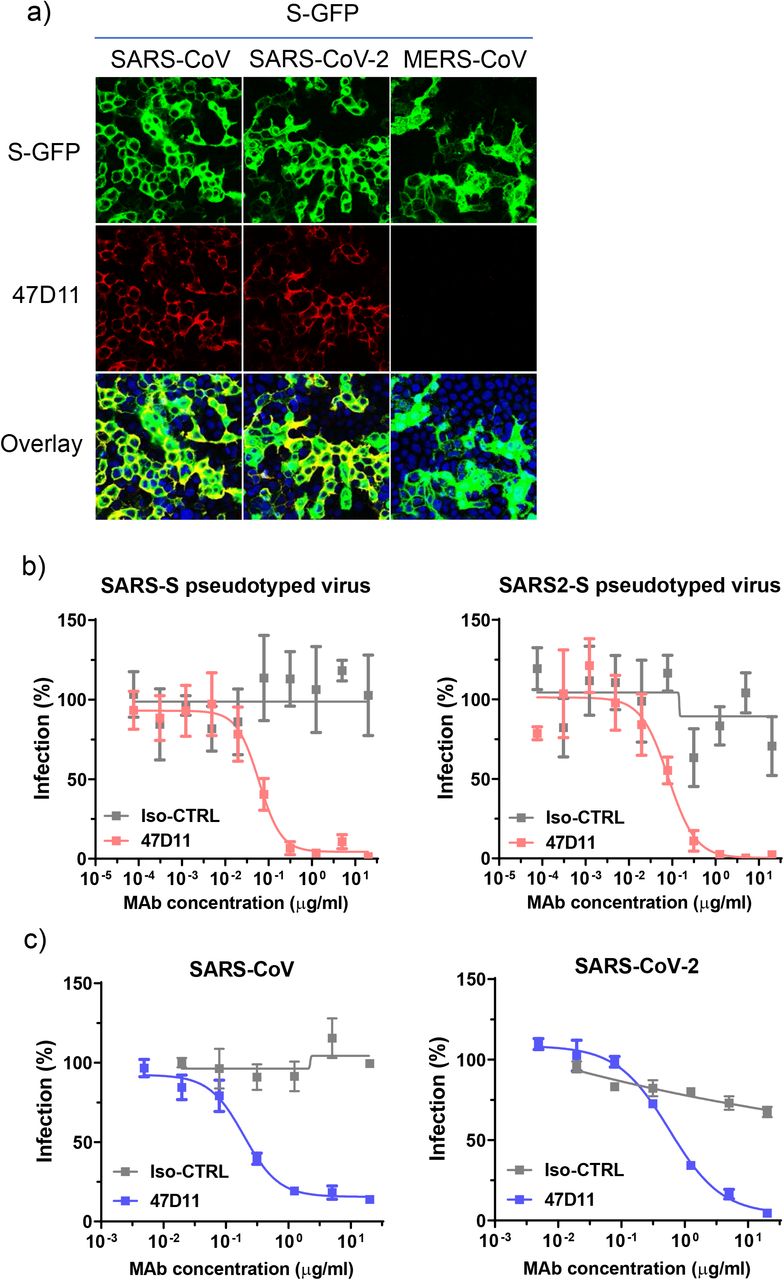
Sl.147D11 neutralizira SARS-CoV i SARS-CoV-2.
a) Vezanje 47D11 na HEK-293T stanice koje eksprimiraju GFP-označene spike proteine SARS-CoV i SARS-CoV-2 detektirane imunofluorescentnim testom. Ljudski mAb 7.7G6 ciljanjem meri-COV S1 Bdomena šiljaka uzeta je kao negativna kontrola, stanične jezgre na prekrivenim slikama su vizualizirane s DAPI. b) Neutralizacijom infekcije luciferarazom VSV čestica koje su posredovane antitijelima pseudotipizirane s spikalnim proteinima SARS-CoV i SARS-CoV-2. Pseudotipizirane VSV čestice prethodno inkubirane antitijelima u naznačenim koncentracijama (vidi metode) korištene su za inficiranje VeroE6 stanicama, a aktivnosti luciferaze u staničnim lizatima određene su 24 sata nakon transdukcije za izračunavanje infekcije (%) u odnosu na kontrole koje nisu liječene antitijelima. Prikazuje se prosjek ± SD iz najmanje dva izvedena neovisna pokusa. Izo-CTRL: nevažna izotipska monoklonska antitijela. c) Neutralizacija posredovana antitijelima SARS-CoV i SARS-CoV-2 infekcije na stanicama VeroE6. Eksperiment je izveden sa trostrukim uzorcima, prikazan je prosjek ± SD.

Slika2. Neutralizirajući monoklonska antitijela 47D11 veže domenu vezanja receptora za SARS-CoV i SARS-CoV-2 šiljke bez šiljaka bez eliminacije interakcije S1 B / ACE2 receptora.
a) ELISA krivulje vezivanja od 47D11 do S ecto (gornja ploča) ili S1 A i S1 B (RBD) (donja ploča) SARS-S i SARS2-S obložene u ekvimolarnim koncentracijama. Prikazuje se prosjek ± SD iz najmanje dva izvedena neovisna pokusa. b) protutijela Interferencija s vezanjem S-S1 B SARS-COV i SARS-COV-2 na površini stanice ACE2-GFP analizirana sa tekućom citometrijom. Prije stanice za vezanje, S1 B se pomiješa s mAb (mAb 47D11, 35F4, 43C6, 7.7G6, u obliku H2L2) sa naznačenim specifičnost mAb: S1 B molarni omjer 8: 1 (v Suppl.Fig.4 postoji opsežna analiza pomoću različitih mAb: S1 Bmolarni omjer je). Stanice su analizirani (ACE2-) GFP ekspresiju (x-os) i S1 B vezanja (y-os). Postoci ćelija s negativnim, pojedinačnim ili dvostrukim pozitivnim vrijednostima prikazani su u svakom kvadrantu. c) Divergencija površinskih ostataka u S1 B SARS-CoV i SARS-CoV-2. Gornja ploča: Struktura SARS-CoV spike proteina S1 B RBD u kompleksu s ljudskim ACE2 receptorom (PDB: 2AJF) 18 . ACE2 (boja pšenice) vizualizira se u obliku vrpce. S1 B jezgrena domena (plava) i poddomena (narančasta) prikazana su u površinskoj prezentaciji pomoću PyMOL i prikazana su istim bojama na linearnom dijagramu vrhovnog proteina, s položajima S1 i S2 podjedinica, S ektodomena (S ecto), označene su S1 domene S1 A-D i transmembranska domena (TM). Donja ploča: Slično kao gornja ploča s površinskim ostacima na S1 B SARS-CoV koji se podudaraju sa SARS-CoV-2 obojenom u bijelu boju.
Domena vezivanja receptora SARS2-S1 B (ostaci 338-506) sastoji se od jezgrene domene i poddomene koja veže receptore (ostaci 438-498) koji se izviru iz antiparalne strukture jezgre domene koja izravno zahvaća receptor. U usporedbi sa jezgrom domene S1 B , identitet proteinske sekvence S1 B receptora koji djeluje na poddomeni SARS-S i SARS2-S je značajno niži (46,7% naspram 86,3%; Suppl.Fig.8 i Fig.2c ). Moćna protutijela za neutralizaciju često ciljaju ovaj poddomena koji veže receptor. Međutim, zbog uobičajenih varijacija ovog poddomena, ta antitijela su često specifična za virus i slabo se vežu i neutraliziraju srodne viruse 16 , 17, Unakrsno reaktivna priroda 47D11 ukazuje na to da će antitijelo vjerovatno ciljati na očuvanu jezgrenu strukturu domene vezivanja S1 B receptora. Vezivanje S1 B za 47D11 dalje od sučelja vezivanja receptora objašnjava njegovu nesposobnost za kompromitiranje interakcije šiljaka i receptora.

Suppl. Slika 8.Usklađivanje proteinske sekvence vezivanja S1 B receptora u (RBD) proteina spike SARS-CoV i SARS-CoV-2 pomoću ClustalW.
Numeriranjem se označava položaj ostataka u cijelom duljini šiljka SARS-CoV (Genbank: AAP13441.1) i SARS-CoV-2 (Genbank: QHD43416.1). Zvezdice (*) označavaju potpuno očuvane ostatke, simbol debelog crijeva (:) označava očuvanje između skupina vrlo sličnih svojstava, a simbol razdoblja (.) Označava očuvanje između skupina slabo sličnih svojstava. Sekvence koje odgovaraju jezgrovitoj domeni S1 B receptora i poddomeni vezivanja receptora obojene su u plavoj i narančastoj boji. Četrnaest ostaci koji su uključeni u vezanje SARS-COV S1 B ljudskog ACE2 su istaknute u sivoj 1 .
Zaključno, ovo je prvo izvješće o (ljudskom) monoklonskom antitijelu koje neutralizira SARS-CoV-2. 47D11 veže sačuvani epitop na domeni vezivanja receptora šiljaka objašnjavajući njegovu sposobnost križnog neutraliziranja SARS-CoV i SARS-CoV-2, koristeći mehanizam neovisan o inhibiciji vezanja receptora. Ovo će antitijelo biti korisno za razvoj testova otkrivanja antigena i seroloških ispitivanja kojima se cilja SARS-CoV-2. Neutralizirajuća antitijela mogu promijeniti tijek infekcije kod inficiranog domaćina koji podržava uklanjanje virusa ili zaštititi neinficiranog domaćina koji je izložen virusu 4 . Dakle, ovo antitijelo nudi potencijal za sprječavanje i / ili liječenje COVID-19, a možda i drugih bolesti u nastajanju kod ljudi uzrokovanih virusima iz Sarbecovirus subgena .
Prilozi autora
BJB je dizajnirao i koordinirao studiju. CW, WL, NMAO, RvH i DD su proveli eksperimente. DD, BLH i BJB nadzirali su dio eksperimenata. Svi autori doprinijeli su predstavljenim interpretacijama i zaključcima. BJB je napisao rukopis, BLH, FJMK, ADMEO i FG sudjelovali su u uređivanju rukopisa.
Materijali i metode
Ekspresija i pročišćavanje proteina koronavirusnih šiljaka
Coronavirus spike ectodomains (S ecto ) SARS-CoV (ostaci 1–1,182; soj CUHK-W1; GenBank: AAP13567,1) i SARS-CoV-2 (ostaci 1–1,213; soj Wuhan-Hu-1; GenBank: QHD43416 .1) su eksprimirane prolazno u stanicama HEK-293T sa motivom trimerizacije C-terminala i Strep-oznakom upotrebom pCAGGS ekspresijskog plazmida. Slično, pCAGGS ekspresijski vektori koji kodiraju S1 ili njezine poddomene SARS-COV (S1, ostaci 1-676, S1 , rezidue 1-302, S1 B , ostaci, 325-533), i SARS-COV-2 (S1, ostaci 1-682; S1 A , ostaci 1-294, S1 B , rezidue 329-538) C-terminalno s oznakom Fc domene humanog ili mišjeg IgG ili strep-privjesak dobiveni su kao što je opisano ranije 19, Rekombinantni proteini su eksprimirani u stanicama HEK-293T i afinitet je pročišćen iz supernatanta kulture zrncima proteina-A sefaroze (GE Healthcare) ili streptaktin zrnca (IBA). Čistoća i cjelovitost svih pročišćenih rekombinantnih proteina provjereni su coomassie obojenim SDS-PAGE.
Stvaranje H2L2 mAbs
Miševi H2L2 bili su sekvencionirani u intervalima od dva tjedna pročišćenim S ectorazličitih COV sljedećim redoslijedom: HCoV-OC43, SARS-CoV, MERS-CoV, HCoV-OC43, SARS-CoV i MERS-CoV. Antigeni su ubrizgani u 20-25 µg / mišu koristeći Stimune Adjuvant (Prionics), svježe pripremljen prema uputama proizvođača za prvo ubrizgavanje, dok je pojačavanje učinjeno pomoću pomoćnog sredstva Ribi (Sigma). Injekcije su rađene supkutano u lijevu i desnu prepone (50 μl) i 100 μl intraperitonealno. Četiri dana nakon posljednje injekcije skupljaju se slezena i limfni čvorovi, a hibridomi nastali standardnom metodom koristeći mijelomsku stanicu mijela SP2/0 (ATCC # CRL-1581) kao fuzijski partner. Hibridomi su pretraženi u antigen-specifičnoj ELISA, a oni odabrani za daljnji razvoj, subklonirani su i proizvedeni u malom obimu (100 ml medija). Za ovu svrhu, hibridomi se uzgajaju u mediju bez seruma i bez proteina za kultiviranje hibridoma (PFHM-II (1X), Gibco) uz dodatak neesencijalnih aminokiselina 100X NEAA, Biowhittaker Lonza, Cat BE13-114E). Protutijela na H2L2 pročišćena su iz supernanata kulture hibridoma uz korištenje afinitetne kromatografije proteina. Pročišćena antitijela su pohranjena na 4 ° C do upotrebe.
Proizvodnja humanog monoklonskog antitijela 47D11
Za rekombinantnu proizvodnju ljudskog mAb, cDNA koja kodira 47D11 H2L2 mAb varijabilne regije teškog i lakog lanca klonirana je u ekspresijske plazmide koji sadrže ljudski IgG1 teški lanac i konstantnu regiju lakog lanca Ig kappa (InvivoGen). Oba plazmida sadrže interleukin-2 signalnu sekvencu kako bi se omogućila učinkovita sekrecija rekombinantnih antitijela. Rekombinantni humani 47D11 mAb i prethodno opisani izotip-kontrola (anti-Streptag mAb) ili 7,7G6 mAb proizvedeni su u stanicama HEK-293T nakon transfekcije s parovima ekspresivnih plazmida IgG1 teškog i lakog lanca prema protokolima InvivoGen. Ljudska antitijela su pročišćena iz supernatanta stanične kulture korištenjem afinitetne kromatografije proteina. Pročišćena antitijela su pohranjena na 4 ° C do upotrebe.
Imunofluorescentna mikroskopija
Vezanje antitijela na proteine stanične šiljke SARS-CoV, SARS-CoV-2 i MERS-CoV izmjereno je imunofluoresencijalnom mikroskopijom. Stanice HEK-293T zasađene na staklenim toboganima su transfektirane plazmidima koji kodiraju SARS-S, SARS2-S ili MERS-S-C-krajnje spojene s proteinima zelene fluorescencije (GFP) koristeći Lipofectamine 2000 (Invitrogen). Dva dana nakon transfekcije, stanice su fiksirane inkubacijom s 2% paraformaldehida u PBS-u 20 minuta na sobnoj temperaturi i obojene za jezgre sa 4,6-diamidino-2-fenilindolom (DAPI). Stanice se zatim inkubiraju s mAbs u koncentraciji od 10 µg / ml, 1 sat na sobnoj temperaturi, nakon čega slijedi inkubacija 45 minuta na sobnoj temperaturi s konusom konzerviranih kozjih antiG humanih protutijela iz Alexa Fluor 594 (Invitrogen, Thermo Fisher Scientific).
Test inhibicije vezivanja receptora temeljenog na protočnoj citometriji
Interferencija antitijela na S1 B vezanje na humani ACE2 receptor na staničnoj površini izmjerena je protočnom citometrijom. HEK-293T stanice su nasađene u gustoći od 2,5 x 10 5 stanica po ml u T75 tikvici. Nakon postizanja konfluencije od 70 do 80%, stanice su transfektirane ekspresijskim plazmidom koji kodira humani ACE2-C-krajnji spoj na GFP koristeći Lipofectamine 2000 (Invitrogen). Dva dana nakon transfekcije, stanice se disociraju otopinom disocijacije stanica (Sigma-aldrich, Merck KGaA; kat. Br. C5914). 2,5 ug / ml humanog Fc označena je SARS-S1 B i SARS2-S1 B preinkubiran sa mAb na navedenu mAb: S1 Bmolarni omjeri tijekom 1 sata na ledu i podvrgnuti protočnoj citometriji. Suspenzije pojedinačnih stanica u FACS puferu su centrifugirane na 400 × g 10 min. Stanice su zatim inkubirane sa S1 B i mAb smjese 1 sat na ledu, nakon čega slijedi inkubiranje s Alexa Fluor 594 konjugiranim anti-humana IgG protutijela (Invitrogen, Thermo Fisher Scientific) tijekom 45 min na sobnoj temperaturi. Stanice su podvrgnute protočnoj citometrijskoj analizi s CytoFLEX protočnim citometrom (Beckman Coulter). Rezultate je analizirao FlowJo (verzija 10).
Ispitivanje neutralizacije pseudotipiziranog virusa
Proizvodnja pseudotipiziranog VSV sa SARS-S i SARS2-S izvedena je kao što je prethodno opisano s nekim prilagodbama 11 . Ukratko, stanice HEK-293T transficirane su pCAGGS ekspresijskim vektorima koji kodiraju SARS-S ili SARS2-S koji nose 28- ili 18-aa citoplazmatski trunak repa. Jednog dana nakon transfekcije stanice su zaražene VSV-G pseudotipiziranim VSVΔG koji nosi krijesnicu ( Photinus pyralis) luciferazni reporterski gen. Dvadeset i četiri sata kasnije, supernatanti koji sadrže SARS-S / SARS2-S pseudotipizirane VSV čestice prikupljeni su i titrirani na stanicama VeroE6 bubrega afričkog zelenog majmuna. U ispitivanju neutralizacije virusa mAbs je serijski razrijeđen u dva puta većoj željenoj krajnjoj koncentraciji u DMEM-u dopunjen 1% fetalnim serumom teleta (Bodinco), 100 U / ml penicilina i 100 µg / ml Streptomicina. Razrijeđeni mAbs se inkubiraju s jednakim volumenom pseudotipiziranih VSV čestica tijekom 1 sata na sobnoj temperaturi, inokuliraju se u spojene VeroE6 slojeve u 96-jažnoj pločici i dalje inkubiraju na 37 ° C tijekom 24 sata. Aktivnost luciferaze je mjerena na Berthold Centro LB 960 pločastom luminometru koristeći D-luciferin kao supstrat (Promega). Postotak infektivnosti izračunat je kao omjer očitanja luciferaze u prisutnosti mAbs normaliziranih na očitanje luciferaze u odsustvu mAb. Pola maksimalne inhibitorne koncentracije (IC)50 ) određene su korištenjem logističke regresije s 4 parametra (GraphPad Prism, verzija 8).
Ispitivanje neutralizacije virusa
Neutralizacija autentičnih SARS-CoV i SARS-CoV-2 izvedena je korištenjem testa za neutralizaciju smanjenja plaka (PRNT) kako je ranije opisano, s nekim izmjenama 20 . Ukratko, mAbs su dvostruko serijski razrijeđeni i miješani sa SARS-CoV ili SARS-CoV-2 tijekom 1 sata. Smjesa je tada dodana u VeroE6 stanice i inkubirana 1 sat, nakon čega su stanice isprane i dalje inkubirane u mediju 8 sati. Stanice su zatim fiksirane i obojene pomoću zečjeg anti-SARS-CoV seruma (Sino Biological) i sekundarnog kozjeg anti-zečjeg IgG (Dako) koji je markiran peroksidazom. Signal je razvijen pomoću precipitata koji formira TMB supstrat (True Blue, KPL), a broj zaraženih stanica po jažici se izračunava pomoću ImmunoSpot® analizatora slike (CTL Europe GmbH). Inhibicijske koncentracije polumaksimalno (IC 50) određene su pomoću 4-parametričke logističke regresije (GraphPad Prism verzija 8).
ELISA analiza vezivanja antitijela na CoV spike antigene
Ploče NUNC Maxisorp (Thermo Scientific) obložene su ekvimolarnim količinama antigena na 4 ° C preko noći. Ploče su isprane tri puta s fosfatnim fiziološkim puferom (PBS) koji je sadržavao 0,05% Tween-20 i blokirane s 3% goveđeg serumskog albumina (BSA) u PBS koji je sadržavao 0,1% Tween-20 na sobnoj temperaturi tijekom 2 sata. Dodana su četverostruka serijska razrijeđenja mAbs počevši od 10 µg / ml (razrijeđena u blokirajućem puferu) i pločice su se inkubirale 1 sat na sobnoj temperaturi. Ploče su isprane tri puta i inkubirane s HRP-konjugiranim kozjim anti-humanim sekundarnim antitijelom (ITK Southern Biotech) razrijeđenim 1: 2000 u blokirajućem puferu 1 sat na sobnoj temperaturi. HRP-konjugirano anti-StrepMAb (IBA, Cat.no: 2-1509-001) protutijelo je upotrijebljeno za potvrđivanje ekvimolarnog premaza Strege-označenih spige antigena. HRP aktivnost izmjerena je na 450 nanometar pomoću tetrametilbenzidinskog supstrata (BioFX) i ELISA čitača ploča (EL-808, Biotek). Pola maksimalne učinkovite koncentracije (EC50 ) vrijednosti vezivanja izračunate su nelinearnom regresijskom analizom na krivuljama vezanja pomoću GraphPad Prism (verzija 8).
Zahvale
Zahvaljujemo dr. Yoshiharu Matsuura (Sveučilište Osaka, Japan) za pružanje VSV-G pseudotipiziranog VSV-G lucferaznog virusa VSVΔG-luc, i Yongle Yang, Michael van der Reijden i Rick Janssens za tehničku podršku. Zahvaljujemo Christianu Drostenu (Charité Universitätsmedizin Berlin, Njemačka) na pružanju virusa SARS-CoV-2. Ova studija je rađena u okviru Nacionalnog centra za jedno zdravlje (NCOH) i Inicijative inovativnih lijekova (IMI) Zoonotic anticipacija i pripremljenost [ZAPI projekt; ugovor o dodjeli bespovratnih sredstava br. 115,76 tisuća]. Miševe koji su korišteni u ovom istraživanju pružila je tvrtka Harbor Antibodies BV, kćeri tvrtke Harbor Biomed ( http://www.harbourbiomed.com ). C. Wang je podržan stipendijom Kineskog vijeća za stipendije (broj datoteke CSC201708620178).
Reference
Napomena: Ovo su preliminarna izvješća koja nisu recenzirana. Ne bi ih se trebalo smatrati zaključnim, povezanim sa zdravljem ili ih se u novinskim medijima navoditi kao potvrđenu informaciju.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}